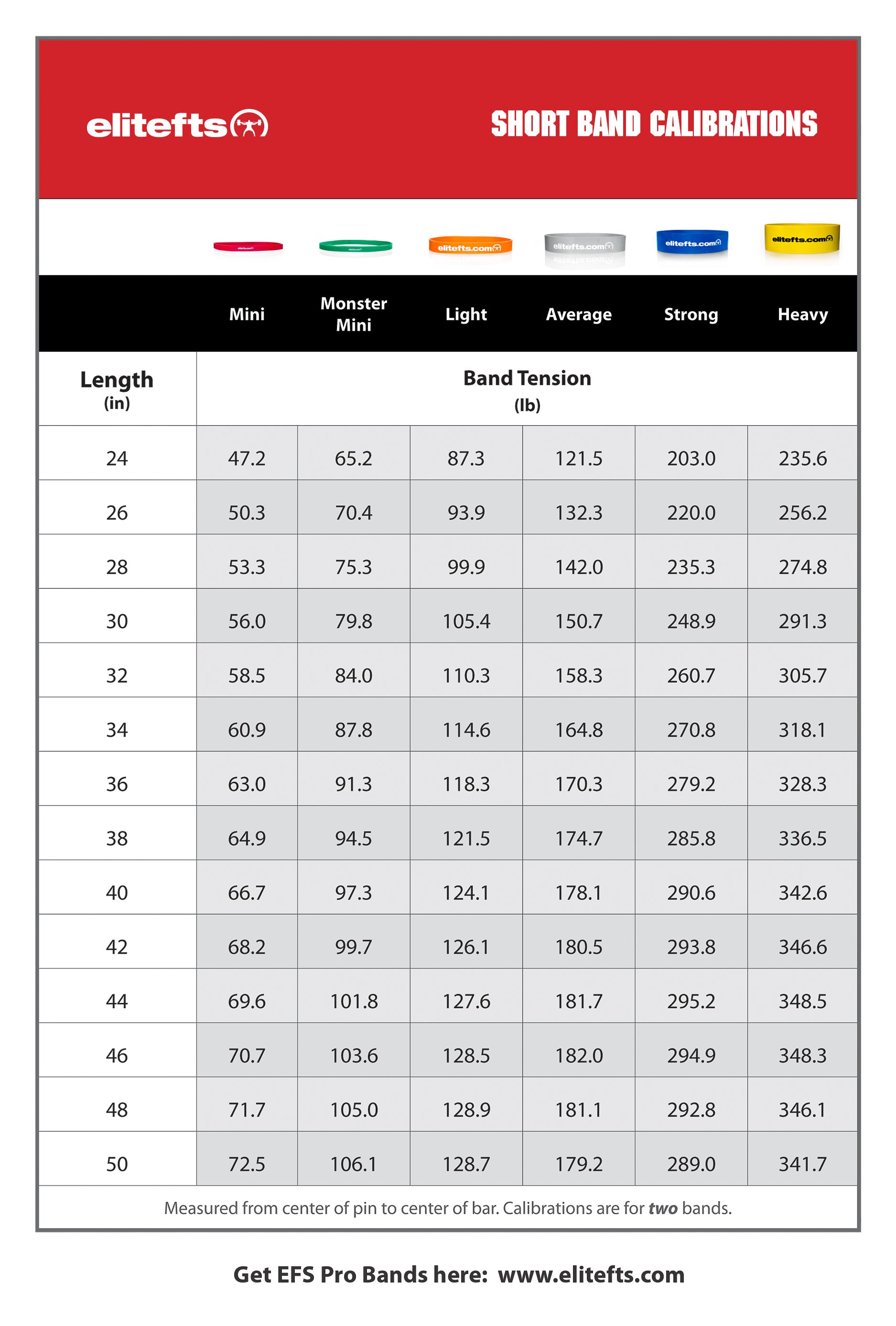
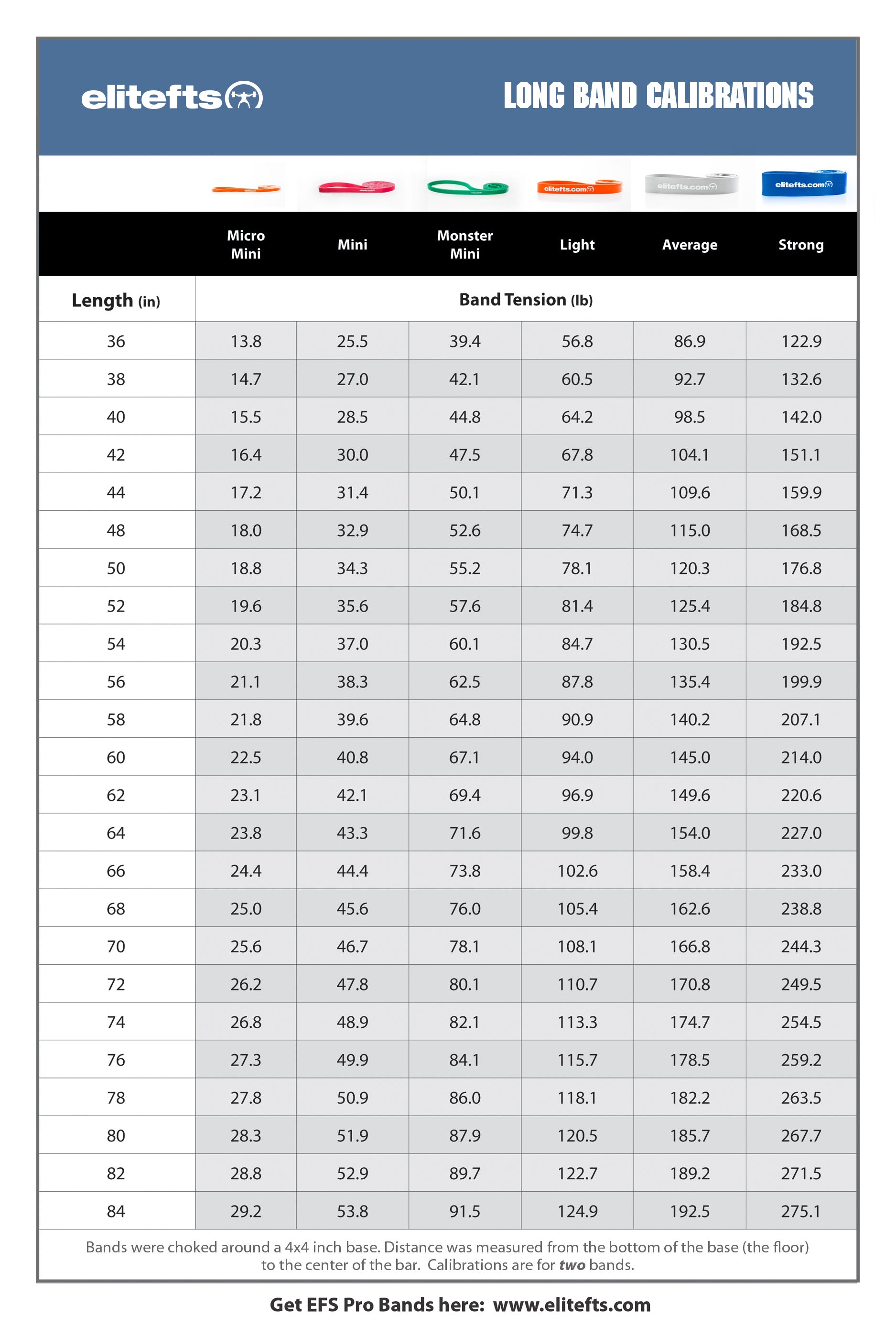
The first article of this series introduced you to a recent quizzical finding that post-exercise myofibrillar protein synthesis (MPS) doesn’t predict muscle growth adaptations to subsequent resistance training(1). Obviously, the process of hypertrophying muscle cells involves increasing contractile protein content(2), but this is an intricate construction project. Conceptually, gross protein synthesis could be considered analogous to the arrival of building materials, in semi-assembled pieces, on the site of a shopping mall construction site. The protein is technically assembled/synthesized, but no one is going to go shopping until there are actual stores up and running. Incorporating this protein, in all the right places and proportions within functional hypertrophied muscle cells entails all the complexity of finalizing the plumbing, electrical, HVAC, roofing, landscaping, etc. that makes that shopping mall such a great temptation to spend too much money.
Indeed, muscle hypertrophy involves the expression of a hundreds of genes(3). To coordinate this construction masterpiece, nuclei are donated via locally enlisted satellite cells, such that the volume of muscle cell under the control of a particular nucleus, its “myonuclear domain size” is maintained(4).
In this article, we’ll see if muscle hypertrophy research using a bigger picture approach, including the importance of satellite cell activity, provides any clues as to why some people grow muscle with relative ease, and others feel like they’re just treading water.
Let’s Get to the Bottom of This
A group of prominent UK researchers used a wide lens molecular biology approach when comparing muscle biopsies before and after training. Subjects’ average hypertrophy after 20 weeks of training ranged from 3 to 28%. This boiled down to the mapping 384 genes and a surprising result. The greatest gains were had in those who had a molecular “signature” characterized by inactive mTOR, i.e., a profile suggesting reduced protein synthesis. Baseline lean muscle mass did not explain who the responders were and neither did sex of the trainee. It was almost universal that the high-responders had this down-regulated mTOR profile(3).
The concluded that, “while acute changes in [mTOR-related] phosphorylation may associate with those of acute remodeling processes (i.e., muscle protein synthesis responses) they are a poor indicator of future lean mass gains(3).” Once again, it seems that simply having a robust protein synthetic response is not all that’s needed when it comes to making good gains. Perhaps an inhibited mTOR profile indicates that slow n’ steady, controlled protein synthesis is a more appropriate strategy for remodeling a larger muscle cell...
Looking Elsewhere
In a study where milk was a better post-workout drink than a macro-matched soy drink [or placebo(5)], members from each supplement condition could be found among in both the bottom, non-responder cluster, as well as the top ~20% of the best responders when it came to muscle growth. More specifically, while overall, milk was a better post-workout recovery supplement, one subject given milk was still among the bottom quartile in muscle growth, while two of the highest responders were consuming the placebo(6). (Does this sound familiar to anyone who has a highly gifted training partner, i.e., one who makes great gains subsisting on “skittles and peanut butter crackers,” while you’re breaking the bank with your food and supplement bill?…)To follow up on this, these researchers analyzed biopsies take before and ~48hr after the last training session and struck gold with a “microRNA” analysis(6). They were able to isolate several microRNAs that specifically differentiated low and high responders. MicroRNAs are ribonucleic acid strands that do not code for a protein, but instead block the translation of protein-coding genes(7). As you can imagine, the interactions here are complex.
Fascinatingly, low responders (but not the subjects who grew the best) had reduced levels of one particular microRNA (miR-378). Research in another context suggests this response might have promoted greater muscle size(8). On the other hand, the “hard gainers” showed a reduction in another microRNA(miR-26a) that could have impaired satellite cell differentiation(9). Once again, these results points to the possibility that “controlled” coordination of growth (as opposed to an all cyclinders firing, maxed-out protein synthetic response), as well as satellite cell involvement (see more below), are both important for packing on the beef.

What About Hormones?
To do justice to the big picture of endogenous hormones and muscle growth would really require an entire article unto itself. The short n’ (not so) sweet of the story is that recent research has basically demonstrated that exercise-induced elevation of (endogenous) hormones like testosterone, IGF-1, and growth hormone is not as important for muscular growth, at least in short term training studies, as once thought(10-12). Ironically, this echoes animal research from more than four decades ago showing that a potent mechanical load can produce tremendous muscle growth, even in the complete absence of these anabolic hormones(13-15).On the other hand, cortisol, perhaps simply because the harder you train, the greater the blood levels(16-19) may predict lean mass gain and (type II) muscle fiber growth(20). More important for our purposes, the extent to which cortisol is suppressed with an intra-workout recovery supplement [that contains carbohydrate(21, 22)] correlates quite well with muscle growth. This has been demonstrated in the same group of subjects in two different studies(23, 24). As I’ve noted before, it could be that (the “sneaky bastard”) cortisol’s slow-acting protelolytic effects(25) keep this glucocorticoid under the immediate post-exercise myofibrillar protein synthesis “radar” (26-28). Indeed, it seems that cortisol may be the most deleterious of the stress hormones(29), easily trumping testosterone’s anabolic effect(30) when it comes to protein breakdown.
The Long Haul
As noted above, growing muscle cells typically increase the number of nuclei they contain, with new genetic material graciously donated by neighboring satellite cells(4). The extent to which this is mandatory for muscle growth has been a topic of “heated” discussion in the scientific community(31-33). It seems that in some instances, satellite cells are not actually needed, especially at the onset of muscle growth(34, 35). This can happen in particular when myostatin levels are suppressed(34, 36), as they just might be after a few workouts (37, 38), but do not seem to be over the long haul of training(39). Perhaps muscle protein synthesis represents an acute alarm reaction, whereas satellite cell activity plays a more important role over the course of larger, more pronounced, long-term increases in cell size.
Indeed, satellite cell behavior seems to have something valuable to tell us about who grows well and who doesn’t. When young trainees show better muscle growth than older ones over months of training(40), the differentiating factor seems to be the ability of satellite cells to do their DNA charity work(41). More relevant to our quandary at hand, over the range from non- to moderate to extreme responders, the ability to bump up muscle nuclei number determines who grows the best. In one study(42), extreme responders blew the other trainees out of the water when it came to turning on myogenin, IGF-1 and mechanogrowth factor(43), all involved in satellite cell activity(44-47). Also, it was only the extreme responders who substantially switched on the production of a particular cell cycle regulating protein (cyclin D1) in the first training session. The authors suggest this is yet another sign that muscle-building Wunderkinder are the best at proliferating (multiplying) satellite cells so that they can donate nuclear material into growing muscle cells(39).
And the Answer Is?...
Several lines of evidence presented in this article suggest that full throttle anabolism, e.g., with the highest rates of MPS or full activation of mTOR, is not associated with the greatest hypertrophic adaptation to resistance training. Within a context whereby some researchers have even suggested the muscle anabolic response is practically unlimited(48), tempering protein synthesis may be part of a hypertrophic strategy that paces MPS with the donation of nuclear material by satellite cells(41, 42). Additionally, some feel that by focusing research efforts MPS, the role of controlling catabolic influences like cortisol(23, 24) may be underestimated when it comes the big anabolic picture(48).Why do some trainees fair better at maintaining the “myonuclear domain” (nucleus / cell volume ratio) and thus end up with greater muscle hypertrophy than others? It seems that the variability may lie in the signaling that coordinates satellite cells activity (see above, e.g., myogenin and microRNAs), not the satellite cells themselves. Unlike rodents(49), humans don’t seem to lose satellite cells over time(50) and the cells themselves retain responsiveness in old age(51, 52). Perhaps the same differences in satellite cell signaling that impair growth in older trainees(41) may also differentiate hard gainers from potential mass monsters(42). Interestingly, the responsiveness of satellite cells seems to vary by muscle(53), which might help explain why some muscle (groups) are simply “stubborn” when it comes to growing.

Satellite Cell Slavery?
Are we slaves to our satellite cells in the quest for muscle size? Of course not, but the concept of “responders” and “hard gainers” does seem to hold some water in the size game(54). You can see it by scanning the landscape in most gyms, as well in analyzing the data in controlled research settings. When it comes to satellite cell recruitment, we know a just little about the particulars of calling them to action. Three sets are better than one(55) for spurring them into action, and low load training both with(56) and without(57) blood flow restriction training gets the job done. Perhaps future research can investigate whether some training parameter [volume, load, frequency(58)] or particular component of the training stimulus [muscle damage(44), progressive tensile overload(59) or metabolic stress(60)] can be exploited to overcome non-responsiveness or age-related growth impairment. It’s also not a stretch to imagine that drugs may be developed to encourage satellite cell activity, e.g., in the aged(61).So, What’s The Take-Home Message?
In circumstances where a particular nutritional intervention improves gains overall(5), there will be variability in responsiveness(6). For every “Bro” who makes great gains doing X, Y and Z, there may be another who does not benefit. From a research standpoint, studies with inbred research animals are great for limiting this source of variation(62). On the other hand, it’s worth considering that animal model (strain) differences, not to mention species (rodent vs. human) add a layer of complexity when trying to apply a study’s finding to the training trenches of real life.For those of you, gym rats and coaches alike, who want to apply bodybuilding related exercise science, recognize too that the very same dataset indicating a given strategy is generally (and statistically) effective may include individual outliers that only the researchers themselves are privy to. Also, be on the lookout for new supplements touting independent effects on satellite cell activity, such as our old friend creatine monohydrate(63-65) and top-selling nitric oxide boosters(66). While it may be true that a given supplement or training technique has demonstrable impact on satellite cell proliferation and/or related growth factors (on average), there may be a more important question: Will said supplement / strategy kick-start satellite cell activity when its severely lacking (the non-responder), or level the playing field for the average Joe who wants to grow like a “genetic freak?”
References
1. Mitchell, C.J., et al., Acute post-exercise myofibrillar protein synthesis is not correlated with resistance training-induced muscle hypertrophy in young men. PLoS One, 2014. 9(2): p. e89431.2. MacDougall, J.D., Morphological changes in human skeletal muscle following strength training and immobilization, in Human muscle power, N.L. Jones, N. McCartney, and A.J. McComas, Editors. 1986, Human Kinetics Publishers: Champaign, IL. p. 269-288.
3. Phillips, B.E., et al., Molecular networks of human muscle adaptation to exercise and age. PLoS Genet, 2013. 9(3): p. e1003389.
4. Van der Meer, S.F., et al., Is the myonuclear domain size fixed? J Musculoskelet Neuronal Interact, 2011. 11(4): p. 286-97.
5. Hartman, J.W., et al., Consumption of fat-free fluid milk after resistance exercise promotes greater lean mass accretion than does consumption of soy or carbohydrate in young, novice, male weightlifters. Am J Clin Nutr, 2007. 86(2): p. 373-81.
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=17684208
6. Davidsen, P.K., et al., High responders to resistance exercise training demonstrate differential regulation of skeletal muscle microRNA expression. J Appl Physiol (1985), 2011. 110(2): p. 309-17.
7. Lewis, B.P., et al., Conserved Seed Pairing, Often Flanked by Adenosines, Indicates that Thousands of Human Genes are MicroRNA Targets. Cell, 2005. 120(1): p. 15-20.
http://www.sciencedirect.com/science/article/pii/S0092867404012607
8. Ganesan, J., et al., MiR-378 controls cardiac hypertrophy by combined repression of mitogen-activated protein kinase pathway factors. Circulation, 2013. 127(21): p. 2097-106.
9. Dey, B.K., et al., miR-26a is required for skeletal muscle differentiation and regeneration in mice. Genes Dev, 2012. 26(19): p. 2180-91.
10. Phillips, S.M., Strength and hypertrophy with resistance training: chasing a hormonal ghost. Eur J Appl Physiol, 2012. 112(5): p. 1981-3; author reply 1985-7.
http://www.ncbi.nlm.nih.gov/pubmed/21898144
11. West, D.W., et al., Elevations in ostensibly anabolic hormones with resistance exercise enhance neither training-induced muscle hypertrophy nor strength of the elbow flexors. Journal of applied physiology, 2010. 108(1): p. 60-7.
http://www.ncbi.nlm.nih.gov/pubmed/19910330
12. West, D.W. and S.M. Phillips, Anabolic processes in human skeletal muscle: restoring the identities of growth hormone and testosterone. The Physician and sportsmedicine, 2010. 38(3): p. 97-104.
http://www.ncbi.nlm.nih.gov/pubmed/20959702
13. Goldberg, A.L., Work-induced growth of skeletal muscle in normal and hypophysectomized rats. The American journal of physiology, 1967. 213(5): p. 1193-8.
http://www.ncbi.nlm.nih.gov/pubmed/6054866
14. Goldberg, A.L., et al., Mechanism of work-induced hypertrophy of skeletal muscle. Medicine and science in sports, 1975. 7(3): p. 185-98.
http://www.ncbi.nlm.nih.gov/pubmed/128681
15. Goldberg, A.L. and H.M. Goodman, Relationship between growth hormone and muscular work in determining muscle size. The Journal of physiology, 1969. 200(3): p. 655-66.
http://www.ncbi.nlm.nih.gov/pubmed/5765853
16. McGuigan, M.R., et al., Salivary cortisol responses and perceived exertion during high intensity and low intensity bouts of resistance exercise. Journal of Sports Science and Medicine, 2004. 3: p. 8-15.
http://jsportscimed.org/vol3/n1/2/v3n1-2pdf.pdf
17. Kraemer, W.J., et al., Hormonal responses to consecutive days of heavy-resistance exercise with or without nutritional supplementation. Journal of Applied Physiology, 1998. 85(4): p. 1544-1555.
http://jap.physiology.org/content/85/4/1544.abstract
18. Crewther, B., et al., The salivary testosterone and cortisol response to three loading schemes. J Strength Cond Res, 2008. 22(1): p. 250-5.
19. Williams, A.G., et al., Effects of resistance exercise volume and nutritional supplementation on anabolic and catabolic hormones. Eur J Appl Physiol, 2002. 86(4): p. 315-21.
20. West, D.W. and S.M. Phillips, Associations of exercise-induced hormone profiles and gains in strength and hypertrophy in a large cohort after weight training. European journal of applied physiology, 2012. 112(7): p. 2693-702.
http://www.ncbi.nlm.nih.gov/pubmed/22105707
21. Ishizuka, B., et al., Pituitary hormone release in response to food ingestion: evidence for neuroendocrine signals from gut to brain. J Clin Endocrinol Metab, 1983. 57(6): p. 1111-6.
22. Volek, J.S., Influence of nutrition on responses to resistance training. Med Sci Sports Exerc, 2004. 36(4): p. 689-96.
23. Bird, S.P., et al., Effects of liquid carbohydrate/essential amino acid ingestion on acute hormonal response during a single bout of resistance exercise in untrained men. Nutrition, 2006. 22(4): p. 367-75.
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=16472979
24. Tarpenning, K.M., et al., Influence of weight training exercise and modification of hormonal response on skeletal muscle growth. J Sci Med Sport, 2001. 4(4): p. 431-46.
25. Simmons, P.S., et al., Increased proteolysis. An effect of increases in plasma cortisol within the physiologic range. J Clin Invest, 1984. 73(2): p. 412-20.
26. Short, K.R., et al., Effect of short-term prednisone use on blood flow, muscle protein metabolism, and function. J Clin Endocrinol Metab, 2004. 89(12): p. 6198-207.
27. Burt, M.G., et al., Impact of Acute and Chronic Low-Dose Glucocorticoids on Protein Metabolism. Journal of Clinical Endocrinology & Metabolism, 2007. 92(10): p. 3923-3929.
http://jcem.endojournals.org/content/92/10/3923.abstract
28. Short, K.R., et al., Short-term prednisone use antagonizes insulin's anabolic effect on muscle protein and glucose metabolism in young healthy people. Am J Physiol Endocrinol Metab, 2009. 297(6): p. E1260-8.
29. Gelfand, R.A., et al., Role of counterregulatory hormones in the catabolic response to stress. J Clin Invest, 1984. 74(6): p. 2238-48.
30. Crowley, M.A. and K.S. Matt, Hormonal regulation of skeletal muscle hypertrophy in rats: the testosterone to cortisol ratio. Eur J Appl Physiol Occup Physiol, 1996. 73(1-2): p. 66-72.
31. O'Connor, R.S. and G.K. Pavlath, Point:Counterpoint: Satellite cell addition is/is not obligatory for skeletal muscle hypertrophy. J Appl Physiol (1985), 2007. 103(3): p. 1099-100.
http://www.ncbi.nlm.nih.gov/pubmed/17289912
32. O'Connor, R.S., et al., Last Word on Point:Counterpoint: Satellite cell addition is/is not obligatory for skeletal muscle hypertrophy. J Appl Physiol (1985), 2007. 103(3): p. 1107.
33. Rehfeldt, C., Satellite cell addition is/is not obligatory for skeletal muscle hypertrophy. Vol. 103. 2007. 1104-1106.
http://jap.physiology.org/jap/103/3/1104.full.pdf
34. Wang, Q. and A.C. McPherron, Myostatin inhibition induces muscle fibre hypertrophy prior to satellite cell activation. J Physiol, 2012. 590(Pt 9): p. 2151-65.
35. McCarthy, J.J., et al., Effective fiber hypertrophy in satellite cell-depleted skeletal muscle. Development, 2011. 138(17): p. 3657-66.
36. Amthor, H., et al., Muscle hypertrophy driven by myostatin blockade does not require stem/precursor-cell activity. Proc Natl Acad Sci U S A, 2009. 106(18): p. 7479-84.
37. Roth, S.M., et al., Myostatin gene expression is reduced in humans with heavy-resistance strength training: a brief communication. Exp Biol Med (Maywood), 2003. 228(6): p. 706-9.
http://www.ncbi.nlm.nih.gov/pubmed/12773702
38. Dalbo, V.J., et al., Acute Loading and Aging Effects on Myostatin Pathway Biomarkers in Human Skeletal Muscle After Three Sequential Bouts of Resistance Exercise. The Journals of Gerontology Series A: Biological Sciences and Medical Sciences, 2011. 66A(8): p. 855-865.
http://biomedgerontology.oxfordjournals.org/content/66A/8/855.abstract
39. Kim, J.S., et al., Resting and load-induced levels of myogenic gene transcripts differ between older adults with demonstrable sarcopenia and young men and women. J Appl Physiol (1985), 2005. 99(6): p. 2149-58.
40. Kosek, D.J. and M.M. Bamman, Modulation of the dystrophin-associated protein complex in response to resistance training in young and older men. Journal of Applied Physiology, 2008. 104(5): p. 1476-1484.
41. Petrella, J.K., et al., Efficacy of myonuclear addition may explain differential myofiber growth among resistance-trained young and older men and women. Am J Physiol Endocrinol Metab, 2006. 291(5): p. E937-46.
42. Petrella, J.K., et al., Potent myofiber hypertrophy during resistance training in humans is associated with satellite cell-mediated myonuclear addition: a cluster analysis. J Appl Physiol (1985), 2008. 104(6): p. 1736-42.
43. Bamman, M.M., et al., Cluster analysis tests the importance of myogenic gene expression during myofiber hypertrophy in humans. J Appl Physiol, 2007. 102(6): p. 2232-9.
44. Hill, M., et al., Muscle satellite (stem) cell activation during local tissue injury and repair. Journal of Anatomy, 2003. 203(1): p. 89-99.
http://dx.doi.org/10.1046/j.1469-7580.2003.00195.x
45. Hill, M. and G. Goldspink, Expression and splicing of the insulin-like growth factor gene in rodent muscle is associated with muscle satellite (stem) cell activation following local tissue damage. The Journal of Physiology, 2003. 549(Pt 2): p. 409-418.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2342958/
46. Le Grand, F. and M.A. Rudnicki, Skeletal muscle satellite cells and adult myogenesis. Current opinion in cell biology, 2007. 19(6): p. 628-633.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2215059/
47. Machida, S. and F.W. Booth, Insulin-like growth factor 1 and muscle growth: implication for satellite cell proliferation. Proc Nutr Soc, 2004. 63(2): p. 337-40.
48. Deutz, N.E. and R.R. Wolfe, Is there a maximal anabolic response to protein intake with a meal? Clin Nutr, 2013. 32(2): p. 309-13.
49. Jejurikar, S.S. and W.M. Kuzon, Jr., Satellite cell depletion in degenerative skeletal muscle. Apoptosis, 2003. 8(6): p. 573-8.
50. Roth, S.M., et al., Skeletal muscle satellite cell populations in healthy young and older men and women. The Anatomical Record, 2000. 260(4): p. 351-358.
http://dx.doi.org/10.1002/1097-0185(200012)260:4<350::AID-AR30>3.0.CO;2-6
51. Renault, V., et al., Skeletal muscle regeneration and the mitotic clock. Exp Gerontol, 2000. 35(6-7): p. 711-9.
52. Mouly, V., et al., The mitotic clock in skeletal muscle regeneration, disease and cell mediated gene therapy. Acta Physiol Scand, 2005. 184(1): p. 3-15.
53. Collins, C.A., et al., Stem cell function, self-renewal, and behavioral heterogeneity of cells from the adult muscle satellite cell niche. Cell, 2005. 122(2): p. 289-301.
54. Timmons, J.A., Variability in training-induced skeletal muscle adaptation. J Appl Physiol, 2011. 110(3): p. 846 - 53.
55. Hanssen, K.E., et al., The effect of strength training volume on satellite cells, myogenic regulatory factors, and growth factors. Scand J Med Sci Sports, 2013. 23(6): p. 728-39.
56. Nielsen, J.L., et al., Proliferation of myogenic stem cells in human skeletal muscle in response to low-load resistance training with blood flow restriction. The Journal of Physiology, 2012. 590(17): p. 4351-4361.
http://jp.physoc.org/content/590/17/4351.abstract
57. Wernbom, M., et al., Acute low-load resistance exercise with and without blood flow restriction increased protein signalling and number of satellite cells in human skeletal muscle. Eur J Appl Physiol, 2013. 113(12): p. 2953-65.
58. Wernbom, M., et al., The influence of frequency, intensity, volume and mode of strength training on whole muscle cross-sectional area in humans. Sports Med, 2007. 37(3): p. 225-64.
http://www.ncbi.nlm.nih.gov/pubmed/17326698
59. Schoenfeld, B.J., The mechanisms of muscle hypertrophy and their application to resistance training. Journal of strength and conditioning research / National Strength & Conditioning Association, 2010. 24(10): p. 2857-72.
http://www.ncbi.nlm.nih.gov/pubmed/20847704
60. Loenneke, J.P., Skeletal muscle hypertrophy: How important is exercise intensity. Journal of Trainology, 2012. 1(2): p. 28-31.
http://trainology.org/PDF/6%20Skeletal%20Muscle%20Hypertrophy%20How%20important%20is%20Exercise%20Intensity.%20Loenneke.pdf
61. Lynch, G.S., Update on emerging drugs for sarcopenia - age-related muscle wasting. Expert Opin Emerg Drugs, 2008. 13(4): p. 655-73.
62. Beck, J.A., et al., Genealogies of mouse inbred strains. Nat Genet, 2000. 24(1): p. 23-25.
http://dx.doi.org/10.1038/71641
63. Olsen, S., et al., Creatine supplementation augments the increase in satellite cell and myonuclei number in human skeletal muscle induced by strength training. J Physiol, 2006. 573(Pt 2): p. 525-34.
64. Hespel, P., et al., Oral creatine supplementation facilitates the rehabilitation of disuse atrophy and alters the expression of muscle myogenic factors in humans. J Physiol, 2001. 536(Pt 2): p. 625-33.
65. Willoughby, D.S. and J.M. Rosene, Effects of oral creatine and resistance training on myogenic regulatory factor expression. Med Sci Sports Exerc, 2003. 35(6): p. 923-9.
66. Shelmadine, B., et al., Effects of 28 days of resistance exercise and consuming a commercially available pre-workout supplement, NO-Shotgun(R), on body composition, muscle strength and mass, markers of satellite cell activation, and clinical sa